Advertisement
Clinical Research and Public HealthInfectious diseaseInflammationMicrobiology
Open Access | ![]() 10.1172/JCI192104
10.1172/JCI192104
Small-molecule correlates of infection precede infection diagnosis in breast implant reconstruction patients
John A. Wildenthal,1 Margaret A. Olsen,1 Hung D. Tran,1 John I. Robinson,1 Terence M. Myckatyn,2 David K. Warren,1 Keith E. Brandt,2 Marissa M. Tenenbaum,2 Joani M. Christensen,2 Thomas H. Tung,2 Justin M. Sacks,2 Rachel A. Anolik,2 Katelin B. Nickel,1 Hideji Fujiwara,1 Peter J. Mucha,3 and Jeffrey P. Henderson1
1Infectious Diseases Division, Department of Medicine and
2Division of Plastic and Reconstructive Surgery, Washington University in St. Louis School of Medicine, St. Louis, Missouri, USA.
3Department of Mathematics, Dartmouth College, Hanover, New Hampshire, USA.
Address correspondence to: Jeffrey P. Henderson, Washington University School of Medicine, 660 S. Euclid Ave., Box 8051, St. Louis, Missouri 63110, USA. Phone: 314.362.7250; Email: hendersonj@wustl.edu.
Find articles by
Wildenthal, J.
in:
PubMed
|
Google Scholar
|

1Infectious Diseases Division, Department of Medicine and
2Division of Plastic and Reconstructive Surgery, Washington University in St. Louis School of Medicine, St. Louis, Missouri, USA.
3Department of Mathematics, Dartmouth College, Hanover, New Hampshire, USA.
Address correspondence to: Jeffrey P. Henderson, Washington University School of Medicine, 660 S. Euclid Ave., Box 8051, St. Louis, Missouri 63110, USA. Phone: 314.362.7250; Email: hendersonj@wustl.edu.
Find articles by Olsen, M. in: PubMed | Google Scholar
1Infectious Diseases Division, Department of Medicine and
2Division of Plastic and Reconstructive Surgery, Washington University in St. Louis School of Medicine, St. Louis, Missouri, USA.
3Department of Mathematics, Dartmouth College, Hanover, New Hampshire, USA.
Address correspondence to: Jeffrey P. Henderson, Washington University School of Medicine, 660 S. Euclid Ave., Box 8051, St. Louis, Missouri 63110, USA. Phone: 314.362.7250; Email: hendersonj@wustl.edu.
Find articles by Tran, H. in: PubMed | Google Scholar
1Infectious Diseases Division, Department of Medicine and
2Division of Plastic and Reconstructive Surgery, Washington University in St. Louis School of Medicine, St. Louis, Missouri, USA.
3Department of Mathematics, Dartmouth College, Hanover, New Hampshire, USA.
Address correspondence to: Jeffrey P. Henderson, Washington University School of Medicine, 660 S. Euclid Ave., Box 8051, St. Louis, Missouri 63110, USA. Phone: 314.362.7250; Email: hendersonj@wustl.edu.
Find articles by
Robinson, J.
in:
PubMed
|
Google Scholar
|
1Infectious Diseases Division, Department of Medicine and
2Division of Plastic and Reconstructive Surgery, Washington University in St. Louis School of Medicine, St. Louis, Missouri, USA.
3Department of Mathematics, Dartmouth College, Hanover, New Hampshire, USA.
Address correspondence to: Jeffrey P. Henderson, Washington University School of Medicine, 660 S. Euclid Ave., Box 8051, St. Louis, Missouri 63110, USA. Phone: 314.362.7250; Email: hendersonj@wustl.edu.
Find articles by Myckatyn, T. in: PubMed | Google Scholar
1Infectious Diseases Division, Department of Medicine and
2Division of Plastic and Reconstructive Surgery, Washington University in St. Louis School of Medicine, St. Louis, Missouri, USA.
3Department of Mathematics, Dartmouth College, Hanover, New Hampshire, USA.
Address correspondence to: Jeffrey P. Henderson, Washington University School of Medicine, 660 S. Euclid Ave., Box 8051, St. Louis, Missouri 63110, USA. Phone: 314.362.7250; Email: hendersonj@wustl.edu.
Find articles by Warren, D. in: PubMed | Google Scholar
1Infectious Diseases Division, Department of Medicine and
2Division of Plastic and Reconstructive Surgery, Washington University in St. Louis School of Medicine, St. Louis, Missouri, USA.
3Department of Mathematics, Dartmouth College, Hanover, New Hampshire, USA.
Address correspondence to: Jeffrey P. Henderson, Washington University School of Medicine, 660 S. Euclid Ave., Box 8051, St. Louis, Missouri 63110, USA. Phone: 314.362.7250; Email: hendersonj@wustl.edu.
Find articles by Brandt, K. in: PubMed | Google Scholar
1Infectious Diseases Division, Department of Medicine and
2Division of Plastic and Reconstructive Surgery, Washington University in St. Louis School of Medicine, St. Louis, Missouri, USA.
3Department of Mathematics, Dartmouth College, Hanover, New Hampshire, USA.
Address correspondence to: Jeffrey P. Henderson, Washington University School of Medicine, 660 S. Euclid Ave., Box 8051, St. Louis, Missouri 63110, USA. Phone: 314.362.7250; Email: hendersonj@wustl.edu.
Find articles by Tenenbaum, M. in: PubMed | Google Scholar
1Infectious Diseases Division, Department of Medicine and
2Division of Plastic and Reconstructive Surgery, Washington University in St. Louis School of Medicine, St. Louis, Missouri, USA.
3Department of Mathematics, Dartmouth College, Hanover, New Hampshire, USA.
Address correspondence to: Jeffrey P. Henderson, Washington University School of Medicine, 660 S. Euclid Ave., Box 8051, St. Louis, Missouri 63110, USA. Phone: 314.362.7250; Email: hendersonj@wustl.edu.
Find articles by Christensen, J. in: PubMed | Google Scholar
1Infectious Diseases Division, Department of Medicine and
2Division of Plastic and Reconstructive Surgery, Washington University in St. Louis School of Medicine, St. Louis, Missouri, USA.
3Department of Mathematics, Dartmouth College, Hanover, New Hampshire, USA.
Address correspondence to: Jeffrey P. Henderson, Washington University School of Medicine, 660 S. Euclid Ave., Box 8051, St. Louis, Missouri 63110, USA. Phone: 314.362.7250; Email: hendersonj@wustl.edu.
Find articles by Tung, T. in: PubMed | Google Scholar
1Infectious Diseases Division, Department of Medicine and
2Division of Plastic and Reconstructive Surgery, Washington University in St. Louis School of Medicine, St. Louis, Missouri, USA.
3Department of Mathematics, Dartmouth College, Hanover, New Hampshire, USA.
Address correspondence to: Jeffrey P. Henderson, Washington University School of Medicine, 660 S. Euclid Ave., Box 8051, St. Louis, Missouri 63110, USA. Phone: 314.362.7250; Email: hendersonj@wustl.edu.
Find articles by Sacks, J. in: PubMed | Google Scholar
1Infectious Diseases Division, Department of Medicine and
2Division of Plastic and Reconstructive Surgery, Washington University in St. Louis School of Medicine, St. Louis, Missouri, USA.
3Department of Mathematics, Dartmouth College, Hanover, New Hampshire, USA.
Address correspondence to: Jeffrey P. Henderson, Washington University School of Medicine, 660 S. Euclid Ave., Box 8051, St. Louis, Missouri 63110, USA. Phone: 314.362.7250; Email: hendersonj@wustl.edu.
Find articles by Anolik, R. in: PubMed | Google Scholar
1Infectious Diseases Division, Department of Medicine and
2Division of Plastic and Reconstructive Surgery, Washington University in St. Louis School of Medicine, St. Louis, Missouri, USA.
3Department of Mathematics, Dartmouth College, Hanover, New Hampshire, USA.
Address correspondence to: Jeffrey P. Henderson, Washington University School of Medicine, 660 S. Euclid Ave., Box 8051, St. Louis, Missouri 63110, USA. Phone: 314.362.7250; Email: hendersonj@wustl.edu.
Find articles by Nickel, K. in: PubMed | Google Scholar
1Infectious Diseases Division, Department of Medicine and
2Division of Plastic and Reconstructive Surgery, Washington University in St. Louis School of Medicine, St. Louis, Missouri, USA.
3Department of Mathematics, Dartmouth College, Hanover, New Hampshire, USA.
Address correspondence to: Jeffrey P. Henderson, Washington University School of Medicine, 660 S. Euclid Ave., Box 8051, St. Louis, Missouri 63110, USA. Phone: 314.362.7250; Email: hendersonj@wustl.edu.
Find articles by Fujiwara, H. in: PubMed | Google Scholar
1Infectious Diseases Division, Department of Medicine and
2Division of Plastic and Reconstructive Surgery, Washington University in St. Louis School of Medicine, St. Louis, Missouri, USA.
3Department of Mathematics, Dartmouth College, Hanover, New Hampshire, USA.
Address correspondence to: Jeffrey P. Henderson, Washington University School of Medicine, 660 S. Euclid Ave., Box 8051, St. Louis, Missouri 63110, USA. Phone: 314.362.7250; Email: hendersonj@wustl.edu.
Find articles by
Mucha, P.
in:
PubMed
|
Google Scholar
|
1Infectious Diseases Division, Department of Medicine and
2Division of Plastic and Reconstructive Surgery, Washington University in St. Louis School of Medicine, St. Louis, Missouri, USA.
3Department of Mathematics, Dartmouth College, Hanover, New Hampshire, USA.
Address correspondence to: Jeffrey P. Henderson, Washington University School of Medicine, 660 S. Euclid Ave., Box 8051, St. Louis, Missouri 63110, USA. Phone: 314.362.7250; Email: hendersonj@wustl.edu.
Find articles by
Henderson, J.
in:
PubMed
|
Google Scholar
|
Published December 23, 2025 - More info
J Clin Invest. 2026;136(4):e192104. https://doi.org/10.1172/JCI192104.
© 2025 Wildenthal et al. This work is licensed under the Creative Commons Attribution 4.0 International License. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
Received: March 27, 2025; Accepted: December 16, 2025
-
Results
Clinical cohort. We enrolled 106 women in 2 distinct clinical cohorts, termed cohorts 1 and 2 (Figure 1). Cohort 1 recruited 50 women from 2018 to 2020 at the time of removal of TEs or permanent breast implants. Since women were recruited at the time of explantation, the reason for implant removal (i.e., infection, noninfectious wound complication, desire to end further reconstruction, or removal at the time of second-stage reconstruction) was already known, and women with clinical implant infection diagnoses were prioritized for recruitment. Cohort 2 recruited 56 women from 2022 to 2023 at the time of mastectomy who were undergoing TE-based reconstruction and were at higher risk of infection (with 1 or more of the following risk factors: obesity, diabetes, or current/recurrent tobacco use). Because women were recruited at the time of mastectomy with implant placement for cohort 2, they were followed longitudinally until the time of TE removal or the sample cutoff date. For both cohorts, implants and breast pocket seroma fluid (if any) were collected by the surgeons in the operating room at the time of implant removal and transported to the laboratory for immediate microbiologic culture workup. Because the seroma fluid specimens were collected and processed identically in the laboratory for the 2 cohorts (see Methods), the metabolomic analyses using the remaining frozen seroma fluids were performed together and results from the 2 cohorts were combined. In cohort 2, additional longitudinal drain fluid samples were collected at routine postmastectomy clinic visits prior to drain removal. Patients were excluded from analysis (n = 36 total) if no seroma fluid was present for metabolomic analysis from the breast pocket at time of implant removal (n = 23), TE removal for second-stage reconstruction was delayed past the sample cutoff date (n = 7), no TE was placed due to a change in clinical management plans (n = 5), or due to revoked consent (n = 1).
Including both cohorts, implants and intraoperative seroma fluid were available from 82 breasts in 70 patients. In total, 36 specimens met our criteria for removal due to infectious causes (see Methods) and 46 specimens for noninfectious causes (34 planned removal for second-stage reconstruction, 10 for noninfectious complications, and 2 from patients with elective removal of the implant contralateral to an infected breast). These 82 intraoperative specimens served as our gold standard samples to assess fluid metabolomes for diagnostic markers, similar to prior studies on breast implant reconstruction infection (40–42). Patient-level demographic information and breast-specific clinical information for the final combined clinical cohort is shown in Table 1 and Table 2, respectively. In addition, across the 44 breasts (32 patients) in cohort 2, we collected 100 longitudinal specimens of fluid approximately weekly from surgical drains placed after mastectomy and immediate reconstruction prior to drain removal. Drain collections past the third clinic visit were excluded due to low numbers.
 Table 1
Table 1Patient-level demographic characteristics for both cohorts of breast implant reconstruction patients
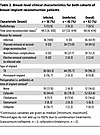 Table 2
Table 2Breast-level clinical characteristics for both cohorts of breast implant reconstruction patients
Bacterial cultures of specimens at time of implant removal. To identify the bacteria most likely to have caused implant infections in our patient population, we cultured seroma fluid samples from the breast pocket and swabbed the explanted implants/TEs for all 82 breasts in our study. All seroma fluids and implants from both cohorts were kept at room temperature until processing for microbial culture, with 79% cultured within 5 hours of specimen collection. A breast was considered culture positive if a bacterium grew from either implant swabs or from seroma fluid. Culture results for the combined cohorts are shown in Table 3. For each bacterial species, recovery in monomicrobial and polymicrobial culture results are shown in Supplemental Table 1 (supplemental material available online with this article; https://doi.org/10.1172/JCI192104DS1). For infected breasts (n = 36), gram-positive bacteria were the most commonly isolated organisms led by Staphylococcus species (n = 16, 44.4%). One or more gram-negative organisms were cultured from 13 (36.1%) infected breasts. The most commonly recovered gram-negative species was Pseudomonas aeruginosa (n = 8, 22.2%). No organisms were cultured from 4 infected breasts (11.1%). Most specimens with Staphylococcus aureus and P. aeruginosa, species with commonly assumed pathogenic roles, were recovered in monomicrobial cultures (Supplemental Table 1). For implants removed at the time of planned second-stage reconstruction in breasts without evidence of infection (n = 34), there was no growth from 20 breasts (58.8%). For implants without evidence of infection that grew 1 or more bacteria (n = 14, 41.2%), skin commensals such as coagulase-negative Staphylococcus (n = 6, 17.6%) and Cutibacterium species (n = 9, 26.5%) were the most common isolates. For implant/TE removal due to noninfectious wound complications (e.g., tissue necrosis and/or dehiscence, n = 10), culture results were more variable and included gram-negative bacteria such as Proteus mirabilis (n = 1, 10%), P. aeruginosa (n = 2, 20%), and Enterobacter species (n = 1, 10%), with no growth in culture of specimens from 5 breasts (50%). Overall, the bacteriology of our clinical cohort was diverse, including both gram-positive and -negative species.
Metabolomic dataset overview. To identify metabolomic markers that differentiate implant infection status, we first used untargeted LC-MS to detect and quantify small molecules in intraoperative seroma fluid stored frozen at –80°C from the combined cohorts. Preprocessing the resulting profiles to minimize duplicate signals and restricting to high-quality features, as described in Methods, yielded 2,540 features for further analysis. Initial hierarchical clustering and heatmap-based analysis did not clearly distinguish infection status (Figure 2A), suggesting that infection was not the predominant driver of metabolomic variation in these samples. To discern infection-associated metabolite features, we used volcano plot analysis (Figure 2B), which revealed 115 features significantly increased at least 4-fold with an FDR-adjusted (Benjamini-Hochberg [BH] method) P (Wilcoxon’s rank sum) < 0.01 in samples from infected breasts. Conversely, 30 features were decreased by at least 4-fold with an FDR-adjusted P < 0.01. To identify features that are robust to variations in physiological dilution, we evaluated pairwise log ratios of feature abundances using differential compositional variation (DCV) scores in the SelEnergyPerm (43) compositional data analysis pipeline. The impact of potential outlier specimens on feature selection was minimized by calculating DCV scores on 25 random 75% subsamples of our dataset and retaining features ranked within the top 20 DCV scores in at least 50% of samplings. This procedure identified 16 features, all of which were increased in infection (Table 4).
 Figure 2
Figure 2Untargeted metabolomic analysis of seroma fluid specimens collected at implant removal. (A) Heatmap of 2,540 features considered for untargeted metabolomic discovery. Color denotes centered and scaled (z score) feature intensity in each sample. (B) Volcano plot of features considered for untargeted metabolomic discovery. The x axis is the mean log2 fold change between specimens collected from infected breasts when compared with specimens collected from breasts with implants removed for noninfectious reasons. The y axis is the adjusted –log10 Wilcoxon’s rank sum P value adjusted for all 2,540 features using the BH method.
 Table 4
Table 4DCV-selected infection-associated metabolomic features in seroma fluid collected at the time of implant removal
Positive molecular correlates of infection. To determine the chemical identities of these 16 infection biomarker candidates, we used accurate mass measurements and LC-MS/MS fragmentation patterns of seroma fluid features to search spectral libraries for matches. This yielded putative matches to acetylated polyamines, peptides, and a glycolipid (Table 4), consistent with metabolites originating in endogenous biochemical processes. The remaining features matched no database entries or were determined to be technical artifacts. Retention time and MS/MS spectrum comparison with reference standards identified the 2 polyamines as N1,N12-diacetylspermine and N1-acetylspermine (Figure 3, A and B, and Supplemental Figure 1). Likewise, the glycolipid matched the retention time and MS/MS spectrum of commercially available glucosyl-sphingosine and was distinguishable from its isomer, galactosyl-sphingosine, a component of myelin (Figure 3C and Supplemental Figure 2). Finally, we putatively identified multiple distinct peptide-like features, including Trp-Glu, Ser-Leu, and Val-Val (Figure 3, D–F), as well as 2 larger peptides (Figure 3, G and H) whose tandem MS/MS spectra contained similarities to di- or tripeptides from spectral libraries (Supplemental Figures 3 and 4). Small peptides may derive from many sources, including de novo production or specific or nonspecific proteolysis from larger polypeptides. Additionally, dipeptides may represent source decay fragments of larger peptides cleaved during ionization in the mass spectrometer and not necessarily the free dipeptide itself. Each identified biomarker candidate exhibited high predictive value, with AUCs of 0.82–0.91 (Table 4).
 Figure 3
Figure 3Seroma fluid features that differentiate infection status. Dot plots and box-and-whisker plots showing prevalence of (A) diacetylspermine, (B) acetylspermine, (C) glucosyl-sphingosine, (D) Trp-Glu, (E) Ser-Leu, (F) Val-Val, and (G and H) 2 unidentified peptide-like features in specimens collected at time of implant removal from uninfected (blue) and infected (red) breasts. Significance is denoted by FDR-adjusted (BH method) Wilcoxon’s rank sum P values (***P < 0.001). Exact FDR-adjusted P values may be found in Table 4. Box-and-whisker plots exclude outliers and depict the minimum and maximum values (whiskers), the upper and lower quartiles, and the median. Dashed lines indicate mean peak area in 5 aligned methanol blanks. As gap filling takes into account sample-specific background, some specimens may have calculated peak areas under this level.
These infection-associated small molecules represent at least 3 categories of endogenously produced biomolecules. Increases in specific acetylated polyamines have been detected in the urine of pulmonary tuberculosis patients (diacetylspermine) and in gram-negative bloodstream infections (N-acetylputrescine and N1,N8-diacetylspermidine) (44–46). Spermidine/spermine acetyltransferase-1 (SAT1), an enzyme that produces N-acetylspermine, is implicated in neutrophil recruitment in a murine tumor model system (47–50). Glucosyl-sphingosine (lyso-GL1, lyso-Gb1) is the deacylated product of glucosylceramides that exhibit antimicrobial activity and are systemically elevated in patients with systemic inflammatory response syndrome (51–54). It may also be a by-product of excess lysosomal lipid degradation (55). While not as widely recognized in this context as cytokines, chemokines, or immune cells, these molecules are robustly associated with infection in the study cohort, implicate distinctive biochemical pathways, and plausibly play roles in immune cell engagement and host defense.
Negative molecular correlates of infection. Although the molecular features that are best able to differentiate infection status were positively associated with infection, several negatively associated features were also detected. In the set of 30 features decreased at least 4-fold with an FDR-adjusted P < 0.01, we identified 2 synthetic molecules: the antibiotic cefazolin (AUC = 0.71; Supplemental Figure 5) and the organic dye crystal violet (AUC = 0.76; Supplemental Figure 6). Eight additional negative correlates were strongly associated with cefazolin (Supplemental Table 2 and Supplemental Figure 5) and likely represent cefazolin metabolites or impurities. Two additional negative correlates were strongly associated with crystal violet (Supplemental Table 3 and Supplemental Figure 6) and had masses consistent with demethylated variants of crystal violet. The negative association between cefazolin and infection corresponds to the preferential use of cefazolin surgical prophylaxis in uninfected compared with infected patients at the time of implant removal (78.3% uninfected vs. 44.4% infected; Table 2). Crystal violet is present in surgical markers used at our institution, and their negative association with infection is attributable to more extensive markings used for planning second-stage reconstructive procedures in uninfected patients. While the post facto anthropogenic origin of these molecules places them outside our goal of tracking biomolecules derived from infection pathophysiology, these findings affirm our overall approach to identifying infection-associated molecules.
Human defensins. A canonical immune response at sites of infection is recruitment of neutrophils and other myeloid cells in response to local stimuli. To quantify this, we sought to measure human neutrophil peptides (HNP1–3; also known as α-defensins 1–3), a series of human myeloid cell defensins that are stored in neutrophil granules prior to release. Qualitative immunoassay-mediated detection of HNPs has been shown to be a marker of infection in prior studies of breast implant reconstruction (40, 41). Although the amphipathic defensins (~3.4 kDa) are larger than the peptides that emerged from our untargeted metabolomic analysis, they are strongly cationic, with multiple charge states that yield ions within the m/z window used for metabolite detection (32). We manually identified LC-MS features in seroma specimens with accurate masses consistent with HNP1–3 that had been filtered out by the peak quality filters (HNP2) or missed by the automated peak caller used for untargeted metabolomic profiling (HNP1 and HNP3). We further confirmed the identify of these distinctive LC-MS features by comparison with commercially available standards (Supplemental Figure 7).
HNP1 (Figure 4A) and HNP2 (Figure 4B) were highly associated with infection (AUCs of 0.93 and 0.92, respectively). The HNP3 AUC was lower (0.78; Figure 4C), likely because 10%–37% of humans lack HNP3 (56). HNP1 and HNP2 levels were highly correlated (Figure 4E), while HNP3 and HNP1 levels exhibited greater deviation (Figure 4F). HNP1 and HNP2 thus exhibit a quantitative association with infection comparable with those for the acetylated polyamines, glucosyl-sphingosine, and other peptide-like features identified above.
 Figure 4
Figure 4Neutrophil presence and activation markers in seroma fluid differentiate infection status. Dot plots and box-and-whisker plots showing prevalence of (A) HNP1, (B) HNP2, (C) HNP3, and (D) 3-chlorotyrosine in specimens collected at time of implant removal from uninfected (blue) and infected (red) breasts. (E) Dot plot showing relative abundance (log10 peak area) of HNP1 versus HNP2. (F) Dot plot showing relative abundance (log10 peak area) of HNP1 versus HNP3. Significance for targeted markers is denoted by unadjusted Wilcoxon’s rank sum P values (***P < 0.001). Box-and-whisker plots exclude outliers and depict the minimum and maximum values (whiskers), the upper and lower quartiles, and the median. For HNP2 and 3-chlorotyrosine, dashed lines indicate mean peak area in 5 aligned methanol blanks. As gap filling takes into account sample-specific background, some specimens may have calculated peak areas under this level. For HNP1 and HNP3 (manually integrated peaks), dashed lines indicate lowest detected abundance in any fluid sample.
Association with neutrophil activation. Because HNPs are prepackaged in cellular vesicles, elevated levels may indicate the presence of myeloid cells but not their degree of activation in response to inflammatory stimuli. To identify neutrophil and monocyte activation, we sought to quantify 3-chlorotyrosine, 3-bromotyrosine, and 3-nitrotyrosine, modified amino acids generated during the respiratory burst when azurophil granules release myeloperoxidase, which generates reactive species through oxidation of chloride, bromide, or nitrite (derived from nitric oxide) ions (57–62). We detected features matching retention times and MS/MS spectra of commercially available reference standards of 3-chlorotyrosine and 3-bromotyrosine (Supplemental Figure 8), but not 3-nitrotyrosine. These features were filtered out by our peak quality filters during initial untargeted analysis. Each was moderately associated with infection status, with AUCs of 0.72 and 0.68 for 3-chlorotyrosine and 3-bromotyrosine, respectively (Figure 4D and Supplemental Figure 8). Of these, 3-chlorotyrosine is the most specific marker of myeloperoxidase activity, which is distinguished from other human peroxidases by its ability to generate chlorinating species (58). 3-Bromotyrosine may be produced by myeloperoxidase or eosinophil peroxidase (59, 63). These results are consistent with a neutrophil cell activation sufficient to stimulate both the respiratory burst and azurophil granule release.
Infection-associated molecules precede infection diagnosis. We hypothesized the existence of a subclinical infectious state preceding implant infection diagnoses. To test this, we measured the infection correlates identified above in longitudinal surgical drain fluid specimens from 44 breasts in 32 patients in cohort 2. Specimens were obtained at the first 3 clinical follow-up visits for 44, 39, and 17 breasts, respectively, with dropouts due to prior drain removal. Visits 1–3 occurred a median of 10, 20, and 27 days after mastectomy with immediate breast implant reconstruction. For the 16 breasts progressing to infection, the timing of drain fluid collection in comparison with implant removal, infection diagnosis, the first recorded instance of erythema, and the first recorded instance of swelling can be found in Figure 5 and Supplemental Table 4. In infected breasts, drain fluid collections on visits 1–3 occurred a median of 36, 29, and 13 days prior to implant removal, respectively. Specimen collections on visits 1–3 preceded infection diagnoses in 16/16, 15/15, and 8/9 breasts (median 35, 23, and 12 days prior, respectively). Notably, 7 of the 16 breasts were only formally diagnosed with infection at the time of implant removal. For infected breasts that developed overt erythema or swelling, specimen collections on visits 1–3 preceded erythema in 8/9, 7/9, and 3/5 breasts (median 28, 18, and 7 days prior to documentation of erythema, respectively) and preceded swelling in 11/11, 10/11, and 3/6 breasts (median 26, 18, and 3.5 days prior to documentation of swelling, respectively). These longitudinal specimens thus sample the postoperative period prior to infection diagnoses and recorded symptom onset.
 Figure 5
Figure 5Timing of clinical events in relation to drain fluid collections. Dot plots and box-and-whisker plots showing the temporal relation of first recorded instance of erythema, first recorded instance of swelling, infection diagnosis, and implant removal in longitudinal drain fluid specimens from breasts in cohort 2 that were removed due to infection (n = 16). (A) First (n = 16), (B) second (n = 15), and (C) third (n = 9) drain fluid collections. All patients had an initial drain fluid collection, but drain removal resulted in dropouts for further time points. Dashed lines at day 0 indicate the date the drain fluid was sampled. Positive values indicate that implant removal, infection diagnosis, first recorded instance of erythema, and/or first recorded instance of swelling occurred after the drain fluid collection. Box-and-whisker plots exclude outliers and depict the minimum and maximum values (whiskers), the upper and lower quartiles, and the median.
At the first postoperative clinic visit, breasts with TEs subsequently removed due to infection exhibited significant (all P < 0.05) but modest (mean 1.7- to 3.9-fold increase, AUC 0.71–0.75) elevations of 3-chlorotyrosine, glucosyl-sphingosine, Trp-Glu, Ser-Leu, and the 556.3 Da peptide-like molecule compared with breasts that did not develop infection (Figure 6 and Table 5). At the second postoperative clinic visit, these differences were sustained for 3-chlorotyrosine (mean 3.0-fold increase), glucosyl-sphingosine (mean 3.0-fold increase), Trp-Glu (mean 7.8-fold increase), Ser-Leu (mean 4.0-fold increase), and the peptide-like molecule at 556.3 Da (mean 6.2-fold increase) and additionally included HNP1 (mean 8.4-fold increase), HNP2 (mean 4.6-fold increase), and Val-Val (mean 6.3-fold increase) with AUCs ranging from 0.77 to 0.83. These markers remained significant at visit 3 (all P < 0.05, mean 4.0- to 21.7-fold increase, AUC 0.81–0.97; Figure 6 and Table 5), although third drain collections were only available for 17/44 breasts. Acetylated spermines and the 1,077.6 Da peptide-like molecule were not consistently different between infected and uninfected breast groups (Supplemental Figure 9 and Table 5). These findings are consistent with the existence of a subclinical infection state preceding implant infection diagnoses, indicated by multiple inflammation-associated molecules detectable in surgical drain fluid.
 Figure 6
Figure 6Infection biomarkers in longitudinal drain fluid collected before implant removal. Dot plots and box-and-whisker plots showing prevalence of (A) HNP1, (B) HNP2, (C) 3-chlorotyrosine, (D) glucosyl-sphingosine, (E) Ser-Leu, (F) Val-Val, (G) Trp-Glu, and (H) peptide-like 556.3 Da biomarkers from longitudinally collected drain fluid in cohort 2 (D1, drain collection 1; D2, drain collection 2; D3, drain collection 3) and seroma fluid collected from breasts at the time of implant removal (SR) in breasts from both cohorts. Specimens are additionally stratified on whether they were from breasts with implants removed due to infection (Inf, red) or remaining uninfected (U, blue). Significance is denoted by FDR-adjusted (BH method) Wilcoxon’s rank sum P values (*P < 0.05, **P < 0.01, ***P < 0.001). Exact FDR-adjusted P values may be found in Table 5. Box-and-whisker plots exclude outliers and depict the minimum and maximum values (whiskers), the upper and lower quartiles, and the median. Dashed lines indicate mean peak area in 5 aligned methanol blanks. As gap filling takes into account sample-specific background, some specimens may have calculated peak areas under this level. For HNP1 (manually integrated), the dashed line indicates lowest detected abundance in any fluid sample.
 Table 5
Table 5Statistical analyses of small molecule biomarkers in longitudinal drain fluid collections and intraoperative seroma fluid
Bacteria preceding infection diagnosis. The presence of increased host-derived infection-associated metabolites in drain fluids from patients who proceed to clinical infection raises the possibility that these patients had subclinical infections at the earlier time points. To assess this possibility, we sought to determine whether the specific bacterial species associated with infection at time of implant removal was also present in preceding drain fluid cultures. The microbiology of drain fluid cultures was diverse (Supplemental Table 5), although P. aeruginosa (n = 17/40 drain collections), coagulase-negative Staphylococcus (n = 10/40 drain collections), and Enterococcus faecalis (n = 11/40 drain collections) were the most common organisms cultured from drain fluid specimens from breasts progressing to infection. Coagulase-negative Staphylococcus (n = 26/60 drain collections) and E. faecalis (n = 12/60 drain collections) were the most common organisms cultured from drain fluid specimens from breasts that did not progress to infection.
Due to the diversity of organisms isolated from drains, we limited our comparison of bacterial presence in drain fluids compared with presence at the time of implant removal to the 9 bacteria isolated from at least 4 separate drain fluid collections (Supplemental Table 6). In cohort 2, 100% (7/7) of breasts that later developed P. aeruginosa infection grew P. aeruginosa in at least 1 preceding drain fluid collection, compared with 2/37 (5.4%) breasts that remained uninfected (n = 1) or had non–P. aeruginosa infections (n = 1) (P < 0.001, 2-tailed Fisher’s exact test). Corynebacterium jeikeium, Serratia marcescens, Staphylococcus warneri, Enterobacter spp., Staphylococcus haemolyticus, S. aureus, E. faecalis, and Staphylococcus epidermidis had no significant association (2-tailed Fisher’s exact test) between presence in at least 1 drain and infection at time of implant removal (Supplemental Table 6). In this cohort, P. aeruginosa recovery from a drain was uniquely associated with subsequent P. aeruginosa breast infection. This is consistent with subclinical infections, though we cannot rule out the possibility that P. aeruginosa in drains is a colonizer, the presence of which predicts subsequent infection.
Bacterial exometabolites precede P. aeruginosa infection diagnosis. The infection metabolome may also include species-specific bacterial exometabolites secreted during infection. Given the high concordance of P. aeruginosa presence in drains and future P. aeruginosa infection, we sought to determine if the well-documented P. aeruginosa exometabolites pyocyanin, dirhamnolipid, monorhamnolipid, and pyochelin (64, 65) were present in the 18 drain specimens positive for P. aeruginosa. These metabolites were identified by retention time and MS/MS spectral matches with authentic standards (Supplemental Figure 10 and Supplemental Table 7). Pyocyanin, P. aeruginosa surfactants (mono- and dirhamnolipids), and the metallophore pyochelin were significantly elevated in drain fluid specimens from which P. aeruginosa was cultured, relative to specimens from which only non–P. aeruginosa species or no bacteria were cultured. The 3 highest intensity metabolites (dirhamnolipid, pyochelin, and pyocyanin) were present in 18/18 (100%) of P. aeruginosa culture-positive drain fluid specimens (Figure 7). These exometabolites were less frequently detected (pyocyanin, 0/9 samples; dirhamnolipid, 4/9 samples; pyochelin, 2/9 samples) in operative seroma fluid specimens that grew P. aeruginosa (8 implants removed due to infection; 1 implant removed due to noninfectious complications) (Figure 7) possibly because the microbial load had been reduced by antibiotic treatment prior to implant removal. Together, these findings suggest that culture-independent P. aeruginosa detection through its distinctive exometabolites may help inform diagnosis and treatment decisions in patients with suspected breast implant–associated infections.
 Figure 7
Figure 7P. aeruginosa metabolites in longitudinal drain fluid and seroma fluid samples. Dot plots and box-and-whisker plots showing prevalence of (A) dirhamnolipid, (B) monorhamnolipid, (C) pyochelin, and (D) pyocyanin in both surgical drains (D) and specimens collected at time of implant removal (S) with specimens that were culture positive (C+) or culture negative (C–). Culture positive samples are further differentiated into samples that did (PA+) or did not (PA–) grow P. aeruginosa. Significance is denoted by unadjusted Wilcoxon’s rank sum P values (**P < 0.01, ***P < 0.001). Box-and-whisker plots exclude outliers and depict the minimum and maximum values (whiskers), the upper and lower quartiles, and the median. Dashed lines indicate mean peak area in 5 aligned methanol blanks. As gap filling takes into account sample-specific background, some specimens may have calculated peak areas under this level.















Copyright © 2026 American Society for Clinical Investigation
ISSN: 0021-9738 (print), 1558-8238 (online)